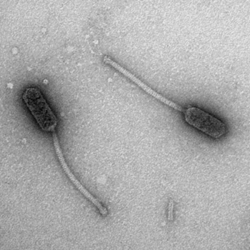
Electron Micrograph of Bacteriophage Babsiella
A virus that infects bacteria.
See 'Phage' for note on use of singular and plural forms.
Also known as: Phage
See also: Mycobacteriophage
Click for more
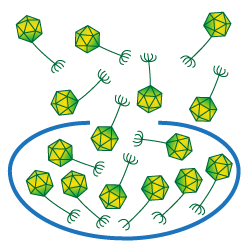
The average number of particles released when a cell is lysed is referred to as a phage's burst size.
The burst size of a phage is the average number of phage particles liberated when an infected bacterium is lysed.
Burst size differs from phage to phage, and can depend on the length of the latent period (the time between infection and lysis). It is likely that burst size has an effect on plaque morphology.
Burst sizes for mycobacteriophages are thought to range from about 50 to 200, though only several phages have been tested.
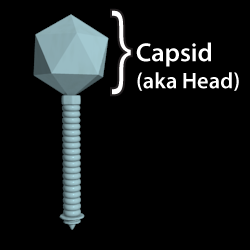
The protein shell (or "head") of a phage. The phage's genetic material is contained within this structure.
Also known as: Head
Click for more
A protein that creates a membrane spanning channel, and aids in the release of phage particles from an infected cell. Frequently found near lysin genes in phage genomes, holins are characterized by transmembrane domains.
Used to refer to a group of phages whose lysogens are immune to infection by other members of the group.
E.g.: Lysogens made by the phage Cjw1 cannot be infected by phage Porky, and vice-versa. Cjw1 and Porky are thus homoimmune.
See also: Lysogen
A recombinase that binds to the AttP and AttB sites between the phage genome and host genome, or AttL and AttR sites in a lysogen. Integrases induce homologous DNA recombination between the two sites to either integrate the phage genome into the bacterial genome or (with the help of an RDF or excise) remove the prophage from the bacterial genome. Mycobacteriophage have two known types of integrases: those that use serine as a catalytic residue, and those that use tyrosine.
A protein that degrades the cell wall and aids in the release of phage particles from an infected cell. These proteins are multi-domain and highly mosaic within the mycobacteriophages. Mycobacteriophages frequently have two lysins: Lysin A and Lysin B.
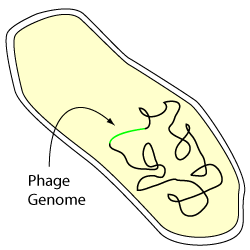
A lysogenic bacterial cell
A lysogen is a bacterium carrying a prophage.
Usually, although not always, the prophage is integrated site-specifically into the host genome although for some phages integration results from transposition. In some examples, the prophage may be propagated extrachromosomally, either as a circular or as a linear genome.
Phage lytic genes are not active in lysogens due to down-regulation by the phage repressor. Lysogens are characteristically immune to superinfection by particles of the same phage type. Phages that share immunity are referred to as being homoimmune. Temperate phages that do not share immunity (i.e. they are capable of infecting lysogens of the other phage) are referred to as being heteroimmune. When grown in lytic growth, lysogens typically release phage particles into the culture supernatent as a result of low levels of spontaneous induction of lytic growth.
Not all phages form lysogens; those that do are called temperate.
See also: Homoimmune, Lysogenic Cycle, Temperate
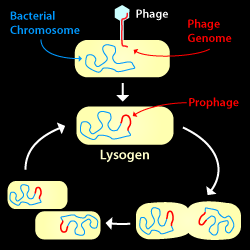
The Lysogenic Cycle: A phage infects a host cell and its genome is integrated into--and propagated along with--the host chromosome.
A phage life cycle where a phage attaches to a bacterium and injects its genome, but does not lyse the host cell. The phage genome is commonly integrated into the host chromosome and replicated along with it when the cell divides. In some cases, however, the phage genome may be propagated outside the host chromosome either in circular or linear form.
Subsequently and under favorable conditions, the prophage may enter the Lytic Cycle, build new virions, and lyse the host cell.
One of two common phage life cycles (the other being the Lytic Cycle).
See also: Lysogen, Lytic Cycle, Temperate
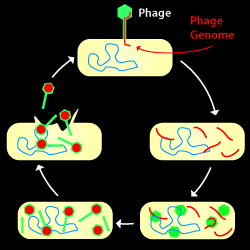
The Lytic Cycle: A phage infects, copies its hereditary material (DNA or RNA), makes new virions, then lyses the cell and escapes.
A phage life cycle where a phage attaches to a host bacterium, injects its hereditary material, takes over the host's machinery, copies its genome, build new virions, then lyses the cell. The offspring produced may then go on to find a new host to infect.
If a phage is strictly lytic (i.e., non-temperate), it generally makes clear (not turbid) plaques.
See also: Lysogenic Cycle
The main component protein of the tail in a flexible tailed bacteriophage. It is assembled around the core of the tail in a spiral pattern with the help of the tail assembly chaperones.
A phage that infects one or more of the Mycobacteria.
See also: Bacteriophage, Phage
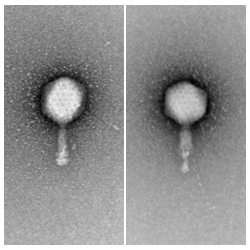
Examples of Myoviridae before (left) and after (right) tail contraction.
A group of phages which have double-stranded DNA genomes and contractile tails.
A small percentage of known mycobacteriophages (~10%, as of early 2011) are myoviridae. These all belong to Cluster C.
See also: Podoviridae, Siphoviridae, Tail
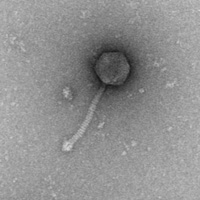
Electron Micrograph of Phage D29
A virus that infects bacteria.
A note on the use of 'phage' and 'phages'. In their book Microbial Genetics by Maloy, Cronan, and Freifelder (Jones and Bartlett; ISBN-10: 0867202483) they comment:
"The plural word ‘phages’ refers to different types of phage, whereas in common usage the word ‘phage’ can be both singular and plural, referring in the plural sense to particles of the same type of phage. Thus P1 and P22 are both phages, but a test tube might contain either 1 P22 phage or 100 P22 phage."
Also known as: Bacteriophage
See also: Mycobacteriophage
Click for more
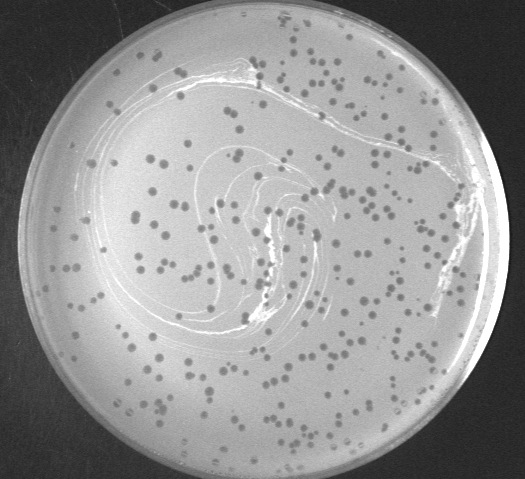
Plaques (zones of death) made by the phage TallGrassMM on a lawn of Mycobacterium smegmatis
An area of clearing in a bacterial lawn where a phage has serially infected and lysed bacterial cells. A "zone of death." Plaques may vary in size or turbidity, but are almost always circular or nearly circular. Agar concentration and moisture content greatly influence plaque size.
Each plaque typically derives from infection of a single viral particle, and the size of plaque increases as the bacteria grow and multiple phage infections occur. Plaque sizes ceases to increase once cells stop growing, although viral particles may continue to diffuse through the top agar layer.
A group of phages which have double-stranded DNA genomes and short, stubby, non-contractile tails.
No known mycobacteriophages are podoviridae.
See also: Myoviridae, Siphoviridae, Tail
The portal is a dodecameric ring made of 12 copies of the portal protein that sits at one vertex of the capsid, replacing one of the capsid pentamers.
During assembly, the terminase binds to the portal and pumps the phage’s genomic material into the capsid. The terminase then dissociates and the tail binds to the portal vertex to form an intact virion.
Earliest detectable stage in capsid assembly. The procapsid is comprised of many copies of the major capsid protein, many copies of the scaffolding protein, the portal, and may include some internal capsid proteins. Procapsids are smaller than the mature capsid and spherical. As the DNA is packaged, the scaffolding protein is removed (either by dissociation or proteolysis) and the procapsid irreversibly expands to the mature icosehedron.
The protease that degrades the scaffolding protein or delta-domain during the transition from procapsid to mature capsid during phage assembly.
A protein that helps the prophage excise from the host genome by binding to the integrase and promoting excision rather than integration. Excise (Xis) proteins, like the ones found in phages lambda and L5, are members of a sub-class of RDFs.
A protein that binds to operators or stoperators within prophage to prevent transcription of lytic genes. Crucial in the maintenance of lysogeny and prophage stability.
A flexible protein that aids in the formation of capsids of the correct size. Scaffolding proteins help form the early stage of the capsid, the procapsid, and then are degraded or dissociate as the capsid expands and matures. They are not present in the mature particle. In some phages, the scaffolding protein is fused to the N-terminus of the major capsid protein. The N-terminal scaffolding domain is then referred to as a delta-domain.
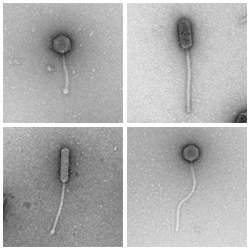
A variety of mycobacteria with a siphoviridae morphology.
A group of phages which have double-stranded DNA genomes and long, flexible, non-contractile tails.
Siphoviridae may have isometric (see, for example, Rosebush) or prolate (see, for example, Babsiella) heads.
The vast majority (~90%, as of early 2011) of known mycobacteriophages are siphoviridae.
See also: Myoviridae, Podoviridae, Tail
Immunity repressor binding sites found in the cluster A mycobacteriophages. These sites are found throughout the entire genome located at the start of many genes. Binding of the immunity repressor during lysogeny to the stoperator sites likely prevents transcription from the entirety of the genome (other than those genes involved in the maintenance of lysogeny). Stoperators differ from standard operators in that they are believed to prevent elongation of transcripts rather than binding of RNA polymerase to promoters.
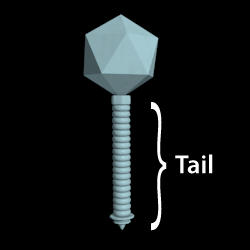
A tube-like protein structure that extends from the phage capsid. Generally, the tip of the tail attaches or adsorbs to the host, and the DNA is injected through the tail into the host cytoplasm.
The type of tails in tailed-phages is used for morphologically-based classification. The Siphoviridae have long flexible non-contractile tails. Myoviridae have contractile tails. Podoviridae have short stubby tails.
See also: Myoviridae, Podoviridae, Siphoviridae, Tapemeasure Protein
Two proteins that help assembly the major tail subunit correctly around the core of the tail. The tail assembly chaperones (called "G" and "G/T" in phage lambda) are frequently located immediately upstream of the tapemeasure in the phage genome. These proteins are translated through a characteristic frameshift.
See also: Tail Chaperone Frameshift
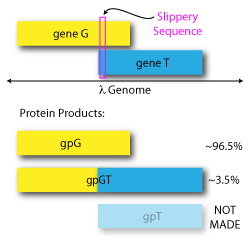
A programmed -1 translational frameshift allows phage λ to make gpG and gpGT in a precise ratio. Similar frameshifts exist in many mycobacteriophages.
A programmed -1 translational frameshift initially observed in bacteriophage Lambda’s tail-assembly chaperone genes.
The frameshift involves two overlapping ORFs, those for gene G and gene T. During translation of gene G, a “slippery” sequence that follows the pattern XXXYYYZ allows the ribosome to occasionally shift backward by 1 nucleotide and continue translation in gene T’s reading frame. When the ribosome does not slip, gpG is made; when the ribosome does slip, a combined product called gpGT is made. The gpT product is not made alone.
Both gpG and gpGT are required for lambda replication, and the ratio between the two is important. In Lambda, gpG is made ~96.5% of the time, and gpGT is made ~3.5% of the time, though this ratio varies in other phages with similar frameshifts.
In mycobacteriophages, a similar frameshift is believed to exist in the tail chaperone genes, although we have been unable to identify the frameshift sequence in phages of Cluster B or Cluster E. In some phages, the frameshift is a +1 shift or -2 shift rather than a -1.
See also: Tail assembly chaperones
Click for more
The instrument of phage attachment to a host cell. Tail fibers are long extended proteins that have the ability to bind and recognize host cells. Fibers are generally very stable: heat, protease, and detergent resistant, and are frequently homotrimers that have coiled-coil or other repetitive secondary structure elements.
This protein is generally encoded by the longest gene in the genome of a flexible tailed phage. It is directly proportional to the length of the tail.
During tail assembly, the tapemeasure joins to the tail tip proteins near its C-terminus, and the tail assembly chaperones bind to tapemeasure long its length to add the major tail subunits. The tail is then capped off and binds to the head.
Tapemeasures may have peptidoglycan degrading motifs near their C-termini to aid in phage infection, and may be injected into the cell during early infection prior to the injection of the phage DNA.
See also: Tail
A temperate phage is one that forms lysogens. Following infection by a temperate phage there are two possible outcomes, lytic growth or lysogeny. Temperate phages therefore form plaques that have turbid centers, due to the growth of lysogens. Turbidity is often more apparent following extended incubation.
See also: Lysogen, Lysogenic Cycle
The ATP-driven motor that packages genetic material into the capsid of the phage. In some phages, the terminase is comprised of two genes: that encoding the large subunit, and that encoding the small subunit. In other phages, only one terminase gene is identifiable. In flexible tailed phages with defined end genomes, the terminase is frequently located at the extreme left end of the genome.